

Зрительные пигменты
Как я уже говорил в начале этой главы, без поглощения вы не сможете видеть. Зрительные пигменты также являются прекрасным примером влияния структуры молекулы на то, как она поглощает свет. И хотя я не могу здесь отдать должное всей обширной и увлекательной литературе по поглощению света фоторецепторами, я хотел бы, по крайней мере, рассмотреть некоторые из основополагающих принципов.
За исключением фитохромов и криптохромов, все фоторецепторы основаны более или менее на одной и той же системе, состоящей из небольшой молекулы, связанной с белком. Молекула, называемая «хромофор», представляет собой каротиноид, производное витамина А. Наиболее часто используемым у обоих позвоночных и беспозвоночных является альдегид витамина А, называемый «ретиналь» (А1). Следующими наиболее распространенными молекулами являются тесно с ним связанные 3,4-дидегидроретиналь, обнаруженный у некоторых рыб, амфибий, рептилий и раков (А2) и 3-гидроксиретиналь (А3), обнаруженных у многих насекомых, в том числе мух и бабочек. У глубоководных кальмаров Watasenia scintillians, помимо A1 и A2, имеется третий хромофор, (4R)-4-гидроксиретиналь (A4).
У каждого из этих четырёх хромофоров немного разные характеристики поглощения. В некоторых случаях несколько хромофоров встречаются у одного и того же животного, иногда одновременно, иногда в разных условиях или на разных этапах жизни. В случае W. scintillians три хромофора, по-видимому, дают ему цветное зрение, что является единственным примером среди головоногих (Seidou et al., 1990). Поскольку большинство животных используют A1, и поскольку основные принципы поглощения света не зависят от типа хромофора, мы остановимся на нём в остальной части этого обсуждения. Мы будем называть его ретиналь, что легче выговорить, чем A1 (и не звучит как номер соуса для стейка).
Ретиналь может быть в двух вариантах: 11-цис и полностью транс. В 11-цис форме молекула перегибается на одиннадцатом углероде; в полностью транс форме молекула является прямой. Поглощение фотона преобразует молекулу назад и вперед между этими двумя состояниями, что похоже на действие тумблера (рисунок 4.13).
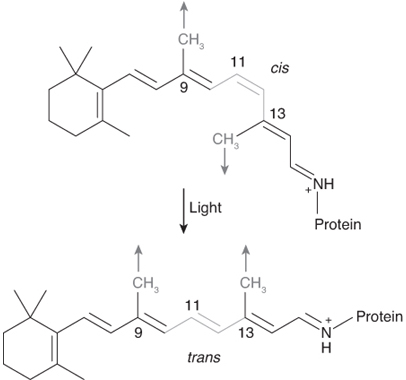
Однако фотоны видимого света не могут это сделать. Фактически, ретиналь наиболее чувствителен к фотонам с длиной волны 340 нм, что значительно в ультрафиолетовой части спектра. Именно здесь вступает в действие вторая часть системы – белок. Хромофор ковалентно связан с большим мембранным белком, известным как «опсин». Опсины являются членами большого сложного семейства белков, известных как «семиспиральные рецепторы, связанные с G-белком», семейство, в которое также входят улавливающие запах молекулы, нейротрансмиттеры и молекулы иммунной системы. В случае улавливания света опсин выполняет две важные функции. Во-первых, когда хромофор меняет форму после поглощения фотона, это изменение заставляет опсин также изменять форму. Изменение формы опсина активирует G-белок внутри клетки, посылающий сообщение о том, что свет был поглощен. В случае позвоночных, опсин теряет свой хромофор после изменения формы и называется «отбеленным» до тех пор, пока не будет прикреплен новый хромофор. У беспозвоночных выпрямленный хромофор остается прикрепленным к опсину и сгибается фотонами различной длины волны. Второе, что делает опсин – это изменение спектра поглощения хромофора. Точно так же, как вжимаясь в сиденье Dodge Dart моего отца я изменял частоту, с которой сиденье вибрировало, попадание хромофора в опсин изменяет длины волн света, поглощающиеся наиболее эффективно.
С эволюционной точки зрения это удобно, поскольку отбор может действовать на ген для производства опсина и изменять спектральную чувствительность зрительной системы в широких пределах. С человеческой точки зрения непросто предсказать спектральную чувствительность заданной комбинации хромофора-опсина исходя из аминокислотной последовательности белка опсина. Мы знаем, что непосредственно окружающие вещество хромофора аминокислоты значат больше всего, но не знаем почему изменение одного остатка с валина на изолейцин сдвигает пик спектра поглощения на 45 нм, а изменение на цистеин сдвигает пик на 3 нм. Это горячая, но в то же время разочаровывающая область исследований, по сложности схожая с предсказанием структуры, в которую сложится белок, исходя из его аминокислотной последовательности.
Очень кстати то, что хотя спектры поглощения зрительных пигментов (то есть комбинаций хромофора-опсина) изменяются в широких пределах, содержа пики, расположенные в диапазоне от 300 до 630 нм, выглядят они все одинаково.
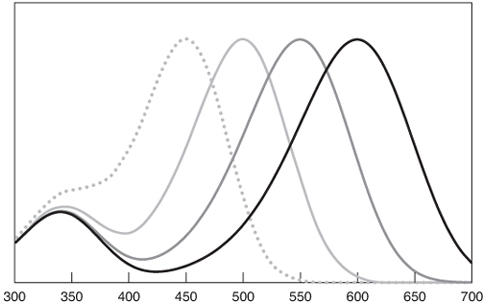
На рис. 4.14 показан набор из четырех визуальных пигментов на основе ретиналя, достигающих пика поглощения на 450 нм, 500 нм, 550 нм и 600 нм. Несколько вещей сразу очевидны. Во-первых, каждая кривая представляет собой сумму двух кривых, вершина одной из которых достигает пика в видимой области, а вершина второй всегда достигает максимума на 340 нм. Пик в видимой области известен как «α-полоса», а в ультрафиолетовой – «β-полоса» соответственно. Кривая β-диапозона почти идентична спектру поглощения изолированного хромофора ретиналя, тогда как α-диапазон изменяется в зависимости от опсина. Если присмотреться, то можно заметить, что α-диапазон немного шире для кривых с пиками на более длинных волнах, но в остальном все они выглядят одинаково. По сути давно известно, что если строить кривую не по длине волны, а по отношению пика длины волны к длине волны (т.е. λmax/λ), вы всегда получаете почти ту же кривую.
Это означает, что вы можете точно предсказать спектр поглощения для зрительного пигмента, если вы знаете две вещи: (1) длину волны, на которой поглощение достигает максимума (известна как «λmax» и произносится «лямбда-макс»), и (2) хромофор, связанный с пигментом. Последнее влияет на форму и пик как α-диапазона, так и β-диапазона. Если это пигмент на основе ретиналя, вам нужно знать только λmax. Существуют мелкие отклонения касательно относительной высоты двух диапазонов, насколько пик β-диапазона зависит от пика α-диапазона и других деталей, но суть в том, что λmax рассказывает вам обо всём, что вам нужно (см. Приложение B для подробных уравнений, основанных на широко используемом шаблоне).
Я понятия не имею, почему это так просто. Вполне разумным кажется ожидать, что некоторые комбинации хромофора-опсина будут иметь совершенно новые распределения спектров поглощения, но никто не нашел их. Это почти похоже на то, что кто-то сознательно упростил жизнь учёным, изучающим зрение.
Однако этим ученым такая услуга может понадобиться, ведь несмотря на то, что кривые поглощения являются простыми и практически идентичными, процесс фототрансдукции (когда начальное изменение хромофора в конечном итоге преобразуется в электрический сигнал) является сложным и варьируется среди таксонов. Опять-таки, на эту тему написана обширная литература и несколько отличных книг (хорошо начать с «Зрение беспозвоночных» (Invertebrate Vision)), поэтому я не буду вдаваться в неё. Тем не менее, остаётся много интересных оптических фактов.
Прежде всего, как я кратко упомянул в главе 2, фоторецепторы подсчитывают фотоны, а не энергию. Ультрафиолетовый фотон 350 нм после поглощения влияет на мембранный потенциал клетки не больше, чем красный фотон 700 нм, хотя первый несёт в два раза больше энергии, чем последний. Спектр поглощения визуального пигмента говорит нам не об эффекте, который будет иметь фотон заданной длины волны, а о вероятность того, что фотон будет поглощен. С точки зрения фоторецепторов все поглощенные фотоны выглядят одинаково.
Во-вторых, клетки-фоторецепторы могут подсчитывать буквально отдельные фотоны. Проводимые с 1950-х годов тщательные эксперименты показали, что клетка-фоторецептор может генерировать электрический отклик на отдельные фотоны, известные как «фотонный щелчок» (обзор Warrant, 2006). Эта чувствительность необычайно велика и соответствует только самым чувствительным из созданных человеком фотометров – фотоэлектронным умножителям. Фоторецепторы не только реагируют на отдельные фотоны, но делают это достаточно надежно. Хотя существуют потери из-за отражения на роговице, поглощения внутри хрусталика и других сред глаза, а также несовершенного преобразования сигналов внутри самого фоторецептора, около половины фотонов, попадающих на роговицу адаптированного к темноте глаза, создают электрический сигнал (обзор по Warrant, 2006).
С учётом этой чувствительности и надежности можно было бы предполагать, что мы сможем хорошо видеть при любой освещённости, но вместо этого наше зрение начинает ухудшаться при освещённости темнее, чем в поздние сумерки, и полностью исчезает на уровне освещённости чуть ниже, чем звёздный свет. Было показано, что человек способен обнаружить вспышку зелёного света объёмом 400 фотонов/см2/секунду (рассмотрен в Warrant, 2006), но любое реальное зрение требует уровней света на пару порядков больше этого.
Для этого есть три основные причины. Первая, и самая главная заключается в том, что независимо от того, как думать о свете – как о волне или как о фотонах – взаимодействие света с веществом происходит в дискретных событиях, подобно тому, как капли дождя попадают в землю. Когда уровень освещенности высок, поток фотонов выглядит довольно непрерывным, как стена июльского ливня. Однако при более низких уровнях освещенности дискретный характер происходящего, известный как дробовой шум или фотонный шум, становится очевидным. Влияние уменьшения освещённости на зрительные способности можно на удивление хорошо оценить количественно, поскольку взаимодействия с фотонами происходят случайным образом, и следуют так называемому «распределению Пуассона». В случайных процессах этого типа, при среднем числе событий, наблюдаемых за данный период времени, равном N, стандартное отклонение этого среднего числа событий будет равно `sqrt( N )`. Таким образом, отношение количества поглощенных фотонов к стандартным отклонениям от среднего составляет N/`sqrt( N ) = sqrt( N )`.
Это отношение, известное как «отношение сигнал-шум» (signal-to-noise ratio,(SNR)) является важным показателем того, насколько хорошо данная зрительная система может видеть при слабом освещении. В частности, оно определяет способность различать два объекта с одинаковой энергетической яркостью, так называемую «контрастную чувствительность». Контрастная чувствительность задается порогом контрастности, процентным отношением разницы в энергетической яркости, которая просто может быть зафиксирована. При ярком свете наш порог контрастности составляет около 1%-2%, в зависимости от типа измерений. По мере уменьшения уровня света этот порог возрастает и обратно пропорционален отношению сигнал-шум. Хорошее эмпирическое правило состоит в том, что порог контрастности Cmin:
4.10
где N - количество поглощенных фотонов на фоторецептор (Warrant, 2006). Число 2.77 не появляется из ниоткуда, но происходит из соотношения между стандартным отклонением среднего и доверительного интервалов. В этом случае порог контрастности устанавливается как процентная разница в энергетической яркости, при которой у вас будет 95%-ный шанс выделить две области различной яркости.
Другими двумя ограничивающими надежность зрения процессами являются тёмный шум и шум приёмника. Тёмный шум относится к тому факту, что иногда фоторецептор будет действовать так, как если бы фотон был поглощен, в то время как он не был. Другими словами, это ложная тревога. Тёмный шум в зрительных пигментах чрезвычайно низок, около 10 событий на фоторецептор в час у насекомых и 360 событий у позвоночных (Warrant, 2006). Он повышается с температурой, поэтому недоказанная гипотеза заключается в том, что глубоководные рыбы (живущие при температуре около 4° C) имеют особенно низкий уровень тёмного шума, что в их случае будет довольно хорошей адаптацией. Шум приёмника является более серьёзной проблемой. Этот шум также относится к физиологии, но в противоположность ложной тревоги он отсылает к тому факту, что электрический отклик на поглощение фотона не является всегда постоянным. Эта случайная вариация при низких уровнях освещенности примерно равна дробовому шуму, поэтому зрение в основном ограничено дробовым шумом и шумом приёмника, тогда как тёмный шум обычно играет второстепенную роль.
Все вместе эти три процесса снижают надежность зрения при низких уровнях освещенности. Множество увлекательных статей было написано о том, как эту проблему обходят ночные и глубоководные животные. Поскольку процессы понижения видимости с падением освещённости хорошо освещены в книгах, посвященных зрению (а также в главе 9, когда мы поговорим об использовании спектрометров), здесь я буду краток. Хотя можно было бы предположить, что ситуации может помочь усиление мощности фоторецептора, эта мера повысит уровень шума так же сильно, как и усилит сигнал. Таким образом, все решения сводятся к одному и тому же – поглощать в фоторецепторе больше фотонов. Добиться этого можно только немногими способами.
Хотя я избегаю использовать в этой книге длинные уравнения, следующее слишком фундаментально и иллюстративно, чтобы им пренебречь. Оно может рассказать вам, сколько фотонов поглощается в отдельном фоторецепторе. Сначала я в общей форме покажу важные для понимания факторы, а затем приведу более конкретную форму, позволяющую рассчитать фактический захват фотонов. В общем виде количество фотонов, поглощаемых фоторецептором, ориентированным на расширенный вид (то есть не на звезду или другой точечный источник света), определяется:
4.11
Давайте обозначим термины. A - площадь зрачка. При прочих равных условиях, чем больше зрачок, тем больше света попадает на каждый фоторецептор. Вот почему крупные телескопы, такие как в обсерватории Кека и Паломарской обсерватории, имеют огромные проёмы; не для увеличения изображения, а для повышения чувствительности. У животных размер зрачка ограничен двумя вещами: (1) большой зрачок требует большого глаза и, видимо, большой головы, и (2) большой зрачок требует большого хрусталика, который трудно создать и, скорее всего, он будет более подвержен помутнению. Таким образом, животное определенного размера может только несколько варьировать A, хотя это и объясняет, почему у ночных и глубоководных животных глаза обычно большие для их размера. Поскольку зрачки обычно круглые, а диаметры (D) легче измерить, чем радиусы, A обычно задается как `( pi D ^2)/4`.
Второй термин Ω - поле зрения (заданное как телесный угол). Единичный фоторецептор поглощает больше фотонов, если он охватываем большую область пространства. Это можно сделать, сделав фоторецептор более широким или сократив фокусное расстояние хрусталика в глазу. Третье решение состоит в том, чтобы связать между собой ряд соседних фоторецепторов, чтобы создать один суперрецептор (рисунок 4.15).

Такая тактика называется «пространственным суммированием». Поскольку она, как представляется, является повсеместной, часто лучше говорить о том, сколько света поглощается неким каналом, где канал может быть либо отдельным фоторецептором, либо набором из них, работающих как один. Все эти решения уменьшают остроту зрения, потому что в системе существенно уменьшается количество пикселей. Однако во многих случаях потеря доли остроты зрения оправдывается усилением чувствительности и, следовательно, надёжности – нет смысла иметь миллионы пикселей, если каждый видит только шум. Например, если матрица физически не становится больше в размерах, увеличение количества пикселей на матрице цифровой камеры снижает её способность работать в тусклом свете. Вот почему иногда покупка 6-мегапиксельной камеры умнее, чем покупка 20-мегапиксельной камеры. Опять же, поскольку люди обычно предпочитают думать в терминах линейных, а не телесных углов, и поскольку поля зрения обычно круговые, то Ω обычно задается как `( pi R ^2)/4`, где R - угловое разрешение канала/рецептора в радианах.
Третий термин Fabs представляет собой долю поступающих в фоторецептор фотонов, которые фактически им поглощаются. Значение зависит от коэффициента поглощения фоторецептора и его длины, и находится с помощью весьма полезного уравнения поглощения 4.8. В этом случае он обычно записывается в виде:
4.12
где l - длина участка фоторецептора, содержащего зрительный пигмент, k - коэффициент поглощения фоторецептора для длины волны с максимальным поглощением, A(λ) – поглощение фоторецептора, нормированное к пику поглощения. Переменные k и A(λ) разделены для удобства. Таким образом, все спектры поглощения имеют пиковое значение, равное единице, и вам просто нужно отслеживать значение k, которые довольно постоянно в какой-либо таксономической группе.
Фоторецепторы уже доверху забиты зрительными пигментами, поэтому самый простой способ увеличить количество поглощаемого света – увеличить длину пути l. Это можно сделать, увеличив длину фоторецептора или пропуская свет через клетку дважды. Последний трюк достигается путем помещения зеркала за фоторецептор, так что свет имеет двойной шанс на поглощение – один раз на входе, и один раз на выходе. Эти зеркала, известные как «тапетум», встречаются в глазах многих животных – посветите фонарём на мордочку вашей кошки, и вы увидите пару примеров. Некоторые животные, особенно глубоководные рыбы, используют как тапетум, так и удлинённые фоторецепторы для поглощения почти каждого фотона, который попадает в глаза. Однако вспомните, что из-за экспоненциального характера передачи света увеличение длины пути в конечном итоге приводит к уменьшению эффективности. Как бы там ни было, в первую очередь никогда не получится уловить больше 100% фотонов, попадающих на рецептор.
Произведение первых трех терминов часто называют «оптической чувствительностью» (s) и оно записывается как:
4.13
Поскольку это почти никогда не упоминается, важно помнить, что оптическая чувствительность зависит от длины волны.
Последним фактором, влияющим на то, сколько фотонов поглощает рецептор, является Δt, время интегрирования. Подобно тому, как камера может контролировать скорость затвора, фоторецептор может контролировать, как долго он помнит о фотоне перед отправкой сигнала. Увеличение времени интегрирования позволяет помнить больше фотонов, поэтому количество дробового шума относительно общего сигнала уменьшается. К сожалению, параллельно также увеличивает количество тёмного шума и шума приёмника, поэтому это не идеальное решение. Другая проблема заключается в том, что увеличение времени интеграции снижает способность обнаруживать быстро движущиеся объекты, так же, как слишком долго открывающийся затвор камеры может размыть фотографию. Время интеграции варьируется в широких пределах: от примерно 0,25 секунды у некоторых медленно движущихся глубоководных животных до менее чем 0,01 секунды у быстролетающих дневных насекомых (Warrant, 2006).
Ещё два последних фактора влияют на захват фотона, но, их значения, как правило, не сильно варьируются. Первый – это передача (τ) хрусталика и других глазных сред (роговица, жидкость стекловидного тела и т. д.). Хотя передача может быть низкой в ультрафиолетовом диапазоне, (если животное имеет УФ-поглощающие пигменты), она обычно относительно высока и постоянна в видимой части спектра (~ 80%). Другим фактором является эффективность квантового преобразования (κ), которая представляет собой долю поглощенных фотонов, фактически вызвавших электрическое событие в клетке. Она была измерена только для нескольких видов, но, по-видимому, относительно постоянна со значением ~ 50%. Объединяя всё это, мы получаем следующее:
4.14
где L(λ) – энергетическая яркость рассматриваемого объекта. Поскольку фоторецепторы не являются спектрометрами и в действительности имеют широкую спектральную чувствительность, обычно вам нужно знать, сколько света было поглощено на всех длинах волн. Таким образом:
4.15
где интегрирование выполняется во всем диапазоне длин волн, и есть измеримое количество света, который способен поглотить конкретный зрительный пигмент (для упрощения я также сложил обычные значения 50% и 80% для эффективности квантового преобразования и передачи). Значение можно ещё уточнять (главным образом за счёт угловой чувствительности фоторецепторов и волноводных эффектов), но уравнение 4.15 даёт хорошую оценку количества поглощаемых фотонов с минимумом возни над подсчётом. Кроме того, как мы обсудим в главе 9, в реальности истинные уровни освещенности редко известны с такой точностью, чтобы волноваться о небольших поправках.
Уравнение 4.15 чрезвычайно полезно, и лежит в основе почти каждого расчета улавливания фотонов, производимого для работ, моделирующих зрительные способности и восприятие цвета у животных. С его помощью можно прогнозировать многие вещи, в том числе контрастную чувствительность и способность отличать цвета друг от друга. Однако нужно быть очень внимательным в отношении единиц. Я видел, как это уравнение используют десятки людей, и могу сказать, что только каждый пятый с первого раза применяет правильные единицы измерения. Самая большая совершаемая ошибка – давать R в градусах, когда она должна быть дана в радианах. Если есть необходимость, R можно изменить на градусы, но не забудьте разделить всё на 3283 (число квадратных градусов в стерадиане). Вторая ошибка заключается в том, что не используются одни и те же единицы как для диаметра зрачка, так и для энергетической яркости. Если энергетическая яркость дана на см2, как это часто бывает, то диаметр зрачка тоже должен быть задан в см. Точно так же время интегрирования должно быть задано в тех же единицах, которые использовались для энергетической яркости, обычно в секундах. Последняя большая ошибка заключается в том, что используются разные единицы для k и l возведённых в степень. Помните, что единицы в возведении в степень отменяются. Итак, если коэффициент поглощения задан в мкм-1, то длину фоторецептора тоже лучше бы дать в мкм. Наконец, полезно помнить, что dλ является реальной единицей, поэтому, если вы делаете интегрирование добавляя столбец чисел в программу электронных таблиц (как делают многие из нас), обязательно умножайте сумму на интервал длины волны. Общий итог для уравнения 4.15 состоит в том, что все единицы должны отменяться, за исключением фотонов.
Как я уже говорил, есть несколько замечательных книг о зрении, более полно раскрывающие тонкие материи преобразования, оптимального пространственного и временного суммирования, теорию волновода фоторецепторов и многие другие увлекательные темы, поэтому здесь я в заключение остановлюсь на одном примере.
Возможно, вы заметили, что я упомянул, что пики поглощения зрительных пигментов колеблются примерно от 300 до 630 нм, и вы, возможно, задавались вопросом, почему не существует зрительных пигментов, пик поглощения которых находится дальше в красном цвете. Это хороший вопрос. На более длинных волнах, конечно, есть много света, по крайней мере, в земных условиях. Также наши ткани ещё достаточно прозрачны для света на этих длинах волн. Закройте фонарик большим пальцем, и вы увидите, что ткани человеческого тела хорошо передают красный свет.
Никто не знает ответа на этот вопрос, но основная гипотеза заключается в том, что зрительные пигменты с пиками на длинах волн более 630 нм слишком чувствительны к тепловому излучению (Ala-Laurila et al., 2004a, 2004b). Поскольку спектры поглощения зрительных пигмента довольно широки, пигмент с пиком 650 нм (например) имеет значительную чувствительность вплоть до 800 нм. Идея заключается в том, что поскольку это достаточно далеко в инфракрасный диапазон, тепловое излучение может вызвать срабатывание фоторецептора. Мне не совсем в это верится, поскольку, как можно видеть из главы 3, количество фотонов 800 нм, создаваемых живыми объектами при физиологических температурах чрезвычайно мало, но на данный момент это лучшая гипотеза из всех.
Ну а что насчёт глубоководных рыб-драконов с красной биолюминесценцией? Производимый ими свет имеет пик на 700 нм. Мы можем конечно увидеть свет с длиной волны 700 нм. На самом деле мы можем видеть даже 800-нм свет. Если вдруг поблизости от вас находится двухфотонный микроскоп, попросите техника осветить лазером с длиной волны 800 нм лист бумаги. Вы увидите красивое красное пятно. Тем не менее, мы можем видеть свет с большой длиной волны только если он очень яркий. Свет от красных фотофоров этих рыбок-драконов тусклый, и как они способны его видеть? Некоторые, как Aristostomias, имеют зрительные пигменты, достигающие максимума на длинных волнах. Тем не менее, другие, подобные Malacosteus, имеют пигмент, который достигает пика только на 542 нм, что недостаточно для того, чтобы увидеть свет 705 нм, который производит само животное.
Удивительно, но похоже, что Malacosteus получает помощь от хлорофилла (Douglas et al., 1998). Как я упоминал ранее, хлорофилл зелёный, потому что он сильно поглощает как синий, так и красный свет. Особенно заметный пик поглощения хлорофилл имеет около 670 нм, близком к пику биолюминесценции Malacosteus. Исходя из этого, и того факта, что хлорофилл известен за его способность передавать энергию другим реакционным центрам, Дуглас и др. (1998) искали и обнаружили в фоторецепторах животного производное хлорофилла, не содержащее магния. Они также обнаружили, что свет 671 нм (близкий к пику поглощения производного хлорофилла) при отбеливании зрительного пигмента действовал лучше, чем свет с длиной волны 654 нм, несмотря на то, что последний был ближе к пику поглощения собственно зрительного пигмента и, следовательно, должен был бы работать лучше при отбеливании.
Гипотеза, развитая в последующих работах (Douglas et al., 1999), состоит в том, что хлорофилл поглощает длинноволновый свет красного фотофора, и передает энергию непосредственно в зрительный пигмент, позволяя последнему функционировать так, как если бы он имел второй пик чувствительности при длине волны 670 нм. Таким образом, хлорофилл выступает как «фоточувствительный пигмент». Хлорофилл животные получают из рациона. Интересно, что Malacosteus, несмотря на то, что имеет зубы и ротовую полость ихтиопода (рыбоядного), в основном питается мелкими планктонными ракообразными, которые содержат хлорофилл, поскольку являются по сути, травоядными (Sutton and Hopkins, 1996).
Наконец, некоторые исследователи считают, что использование хлорофилла для усиления чувствительности к свету с большой длиной волны может быть более распространенным, чем предполагалось ранее, отмечая присутствие хлорофилла у саламандр (Isayama et al., 2006). Другая группа обнаружила, что мыши, инъецированные производным хлорофилла e6 повысили чувствительность к красному свету примерно в два раза (Washington et al., 2007). Зная, что хлорофилл и родопсин являются двумя великими поглощающими молекулами жизни, наблюдение за тем, как они работают вместе для усиления зрительных способностей животных в глубоком море (где видеть непросто, а фотосинтеза не существует) – является прекрасным примером эволюционной экономии и изобретательности природы. Это также достойный способ закончить наше обсуждение поглощения.